Jian Li1,2,3,
Fei Liu3,
Yang Liu4,
Jia-Jun Du1,2,
Qi Liu1 ![]()
For correspondence:- Qi Liu Email: qiliu6655@gmail.com Tel:+8602165161782
Received: 22 May 2015 Accepted: 10 December 2015 Published: 29 January 2016
Citation: Li J, Liu F, Liu Y, Du J, Liu Q. Wortmannin as targeted therapeutic agent for the treatment of triple-negative breast cancer. Trop J Pharm Res 2016; 15(1):95-99 doi: 10.4314/tjpr.v15i1.13
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose:To investigate the inhibitory effect of wortmannin on inhibition of BT-20 and BT-474 breast cancer cells in Athymic nu/nu mice model.
Methods:Forty Athymic nu/nu mice were randomly assigned to 4 groups of 10 each, namely, BT-20 control, BT-20 treatment, BT-474 control and BT-474 treatment groups. The mice were injected with 2.5 x 105 BT-20 and BT-474 cells under anesthesia. Those in the treatment groups were given wortmannin (7 mg/kg) in DMSO daily intraperitoneally whereas the animals in control groups received an equal volume of DMSO for 21 days after the cancer attained palpable stage. Western blot analysis was carried out using enhanced chemiluminescence reagent while Protean IEF cell unit was used for 1-D electrophoresis.
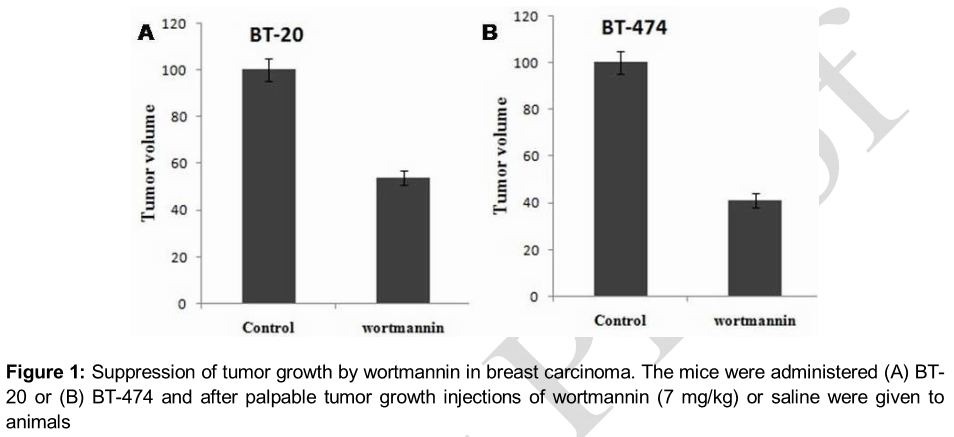
Results:The results showed a significant decrease in the growth of the tested cancer cell lines on treatment with wortmannin at 7 mg/kg daily for 21 days. The volume of tumor in the treatment group was reduced to 42.72 ± 9.45 compared to 79.43 ± 17.11 mm3 in the control group after 21 days treatment (p < 0.005). It also changed the ex
Conclusion:Wortmannin can be of benefit in the treatment of human breast cancer.
Introduction
Breast carcinoma is a challenging health issue at present affecting one out of every eight women, especially in US [1]. It is estimated that about 20 % of the patients with primary breast cancer rapidly develop distant metastases leading to higher mortality rates [2]. The median survival period for breast cancer patients is around three years [2]. Breast cancer has been found commonly in women and its proportion in men is very low [3]. Triple negative breast cancer (TNBC) comprises a group of heterogeneous diseases which are prevalent among African and American women and is more aggressive compared to other cancers [4-6]. It has been reported that TNBC spreads rapidly to distant tissues and leads to mortality within very short time span of the disease [4]. Therefore, the discovery of molecules which target the TNBC is desired for its treatment.
The commonly used chemotherapeutic agents for cancer treatment are antibiotics belonging to the anthracycline, bleomycin, actinomycin, mitomycin and aureolic acid families [7]. Among these families, anthracyclines exhibit promising anti-tumor activities through the suppression of topoisomerase II [8,9]. Another important natural isolate wortmannin from the fungus Talaromyces wortmanni has been shown to act as inhibitor of signal transduction pathways targeting 3 kinase (PI3K) activity [10]. Targeting signal transduction cascades by involvement of bioactive compounds is believed to be a promising strategy for carcinoma therapy [11]. The present study was designed to investigate the effect of wortmannin on breast carcinoma inhibition, miR expression and proteome expression.
Methods
Materials
Cell lines and culture
Human breast carcinoma cell lines, BT-20 and BT-474 were purchased from the Riken Bioresource Center (Tsukuba, Japan). The cells were cultured in cultured Dulbecco's modified Eagle's medium/F-12 (DMEM/F-12) (Wako) supplemented with 10 % fetal bovine serum (FBS) (Cambrex, Walkersville, MD, USA).
Drug
Wortmannin was isolated from the fungus, Talaromyces wortmanni, using solvent extraction followed by column chromatography in 10 % methanol and chloroform.
Xenograft model of tumorigenesis
Athymic nu/nu 6-week-old female mice (Harlan Sprague-Dawley) were fed irradiated sterile food and water. The mice were acclimatized to the laboratory environment one week before the experiment was started. The animals were randomly assigned to 4 groups with 10 each, BT-20 control, BT-20 treatment, BT-474 control and BT-474 treatment groups. The animals were injected with BT-20 or BT-474 cells (2.5 x 105) in the mammary fat pad under anesthesia. The animals in the treatment groups were given wortmannin (7 mg/kg) in DMSO daily intraperitoneally whereas the animals in control groups received equal volume of DMSO for 21 days after the cancer attained palpable stage. During the treatment period volume of cancerous mass was calculated after every 2 days by measuring the tumor size with digital calipers. After the completion of the treatment, the animals were sacrificed to extract the tumors, uterus, liver, and lungs. The tissues were immediately fixed in formalin and stored under cold conditions using liquid nitrogen. The liquid nitrogen frozen samples were then used for other analyses.
The animal experiments were performed according to the rules of the ethical committee of the Weifang People’s Hospital, No. 207 Guangwen Street, Kuiwen District, Weifang, Shandong. All measures were taken to minimize suffering in the animal.
Western blot analysis
The cells after treatment with wortmannin were washed with phosphate-buffered saline (PBS) and lysed in lysis buffer containing aprotinin (10 mg/mL), leupeptin (10 mg/mL) (ICN Biomedicals, Inc., Asse Relegem, Belgium) and phenylmethylsulfonyl fluoride (Sigma-Aldrich). The proteins were separated on SDS-PAGE, loaded on PVDF and treated with primary antibodies. After incubation the membrane was incubated with secondary antibodies conjugated to horseradish peroxidase. The blots were examined using enhanced chemiluminescence reagent (ELPS, Seoul, Korea).
Proteomics analysis
The cells, after treatment with wortmannin or normal saline for 24 h, were subjected to 2D electrophoresis. Protean IEF cell unit (BioRad, Hercules, CA, USA) was used for 1-D electrophoresis and the proteins were separated using Precast 11 cm IPG strips. BioRad Criterion electrophoresis cell system was employed for 2-D electrophoresis and Gel Doc XR image system (BioRad) was used for analysis of stained gels. The scanned gels were analyzed with PDQuest software (version 8.01).
Statistical analysis
All the experiments were performed in triplicate. Student t-test or analysis of variance (ANOVA) was used for statistical analysis. Differences were considered statistically significant at p < 0.05.
Results
Inhibition of breast carcinoma growth
The animals in both treatment and control groups were administered carcinoma cells, BT-20 or BT-474 along with reduced growth factor matrigel (A,B). When the tumor attained palpable stage, animals in the treatment group were given wortmannin (7 mg/kg/day) and those of control group received an equal volume of normal saline. It was observed that the volume of tumor in treatment group was markedly small compared to control group. The tumor volume in the treatment and control groups were 42.72 ± 9.45 and 79.43 ± 17.11 mm3, respectively after the completion of the treatment period.
Effect of wortmannin on the miRnome
The BT-20 cells on exposure to wortmannin for 24 h showed prominent changes in the expression of miRNA whereas no such changes were observed in untreated control cells. Wortmannin treatment significantly enhanced the expression of some of the miRs including miR 19a/b, 22, 29b/c, 181c/d, 195 and 663 (A). In addition wortmannin treatment inhibited the expression of some of the miRs including, 193a 5p, 197, 224, 486 5p, and 542 5p (B).
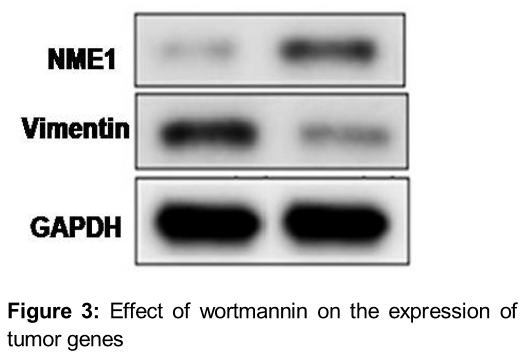
Wortmannin changes the proteome of BT-20 breast carcinoma cells
Exposure of BT-20 cells to wortmannin for 24 h induced changes in the expression of proteomes. The results from Western blot analysis showed increase in the expression of NME1 (NM23 H1) by wortmannin treatment (). The expression of vimentin was significantly decreased by treatment with wortmannin ().
Discussion
In cancerous tissues miR expression shows changes which are known to be associated with increase in tumor growth, resistance to drugs and spread of cancer cells [16-18]. The results obtained in the present study showed that wortmannin induces expression of miRs and thus acts as suppresser of tumor growth. It has been reported that miRs exhibits anti-carcinoma effect on tumor growth by interfering with the expression of Jun B and Jun D [19]. miR 19 suppresses the expression of tissue factor which is involved in the progress of tumors [20]. The results from the present study revealed increase in the expression of tumor suppressor genes on exposure to wortmannin. Studies have demonstrated that miR 193a 5p genes play an important role in the progression of tumor growth [21]. In the present study it was observed that the expression of the genes was inhibited by exposure to wortmannin.
It is reported that miR 21 gene is responsible for inducing resistance to drugs resulting in poor tumor prognosis rate [22,23]. Wortmannin induced inhibition of these genes and thus prevented the development of resistance to drugs. A surface marker found during change of epithelial cells to mesenchymal cells is over-expressed in breast cancer cells [15]. The expression of vimentin is significantly enhanced following treatment with wortmannin. Therefore, wortmannin suppresses the growth of tumors in breast cancer cells.
Conclusion
Wortmannin inhibits the growth of breast carcinoma cells via change in expression of miR. Thus, wortmannin can be used for the treatment of breast cancer.
References
Archives


News Updates